
Zytoskelett In fluoreszenzmikroskopischen Bildern mag das Zellskelett tatsächlich wie ein Skelett aussehen, der Name ist aber irreführend, weil es sich beim Zytoskelett nicht um ein steifes Skelett oder Gerüst handelt, sondern um ein dynamisches Geflecht von Strukturen. Man weiß inzwischen auch, dass Zytoskelettelemente nicht nur für die mechanische Stabilität, sondern auch für sensorische Funktionen wie die Signalübertragung zwischen Zellen unerlässlich sind. Das Wort Zytoskelett wurde 1931 vom Embryologen Paul Wintrebert geprägt, der darüber spekulierte, wie es Eizellen gelingen könnte, ihre interne Organisation zu erhalten, wenn sie durch den engen weiblichen reduktiven Trakt wandern müssen. Aus den starken auf die Eizelle einwirkenden Deformationskräften leitet Wintrebert die Notwendigkeit der Existenz eines Zytoskeletts („cytosquelette“) ab.[2] Das eukaryotische Zytoskelett[Bearbeiten] Endothelzellen unter dem Mikroskop. Mikrotubuli[Bearbeiten] Aktinfilamente[Bearbeiten] Weblinks[Bearbeiten]
Organite Un article de Wikipédia, l'encyclopédie libre. Principaux organites[modifier | modifier le code] Le réticulum endoplasmique (5 et 8), constitué d'un ensemble de replis et de tubules membranaires qui délimitent un compartiment interne séparé du reste du cytosol. Certaines protéines, lors de leur synthèse, transitent par le réticulum pour y être maturées. Organites présents chez les cellules végétales et les champignons[modifier | modifier le code] La vacuole, délimitée par une membrane simple (appelée tonoplaste) assure des fonctions de dégradation cellulaire (comme les lysosomes des cellules animales), de stockage (protéines, sels, substances toxiques, etc.), d'occupation de l'espace (dans des grandes cellules végétales qui se gonflent d'eau par osmose). Organites spécifiques de la cellule végétale[modifier | modifier le code] Les plastes, contenant leur propre génome. Autres structures cellulaires[modifier | modifier le code] Histoire et terminologie[modifier | modifier le code]
Aktin G-Aktin (PDB code: 1j6z) mit ADP und Kation F-Aktin; Darstellung von 13 Aktin-Untereinheiten (basierend auf dem Filamentmodell von Ken Holmes [1]) Geschichte[Bearbeiten] Aktin wurde erstmals 1887 von William Dobinson Halliburton experimentell aufgezeigt.[3] Er untersuchte in Analogie zur Gerinnung des Blutplasmas die Bedingungen, unter denen Proteine in Zellflüssigkeiten der Muskulatur ihre Zustandsform verändern („koagulieren“), und stellte den Einfluss eines Extraktes heraus, das er als „Myosin-Ferment“ bezeichnete.[4] Halliburton konnte seine Forschungen nicht weiter vertiefen, sodass die eigentliche Entdeckung des Aktins heute Brunó Ferenc Straub zugeschrieben wird, der als Forschungsassistent im Labor Albert Szent-Györgyis am Institut für medizinische Chemie der Universität von Szeged in Ungarn über Proteine in Muskelzellen arbeitete. Auf Grund der Kampfhandlungen während des 2. Die Abfolge der Aminosäuren in der Polypeptid-Kette eines Aktins wurde 1973 von M. Aufbau[Bearbeiten]
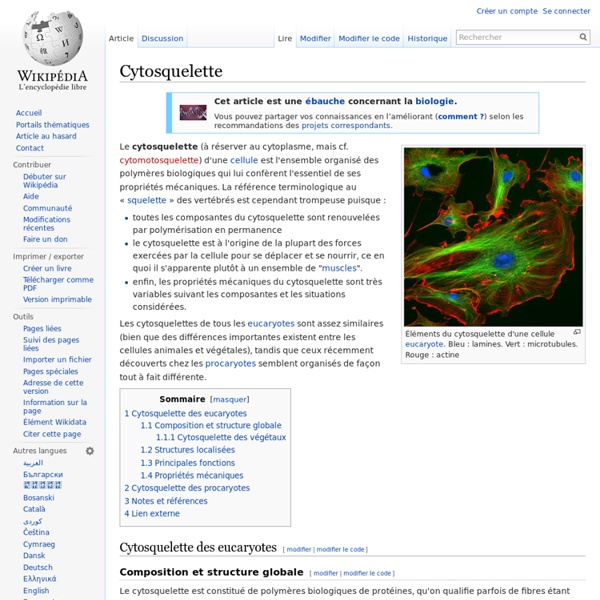
Actine Un article de Wikipédia, l'encyclopédie libre. Actine G Actine F L'actine est une protéine bi-globulaire de 5,46 nm de diamètre importante pour l'architecture et les mouvements cellulaires. Elle est présente dans toutes les cellules du corps (protéine ubiquitaire), et spécialement dans les cellules musculaires. Elle peut représenter jusqu'à 15 % de la masse totale protéique des cellules. Structure[modifier | modifier le code] Chez les mammifères, il existe 6 isotypes d'actine : 3 isoformes d’actines alpha (présentes dans les muscles striés squelettiques et cardiaque et les muscles lisses),2 isoformes d’actines gamma (présentes dans le muscle lisse entérique et dans les tissus non musculaires),1 isoforme d’actine beta (non musculaire). L'actine est une protéine dont le diamètre est de 5,5 nm, qui contient un acide aminé rare : la 3-méthyl histidine. Actine G (globulaire), forme monomérique soluble en solution aqueuse. Polymérisation[modifier | modifier le code] Autres rôles :
Organell Schemazeichnung einer tierischen Zelle. Das Cytoplasma (11) wird nicht als Organell bezeichnet. Einzellige Lebewesen ohne Zellkern (Prokaryoten) haben in der Regel keine Membranen im Inneren der Zelle und demnach auch keine Organellen nach der ersten Definition. Es gibt jedoch prokaryotische Strukturen, die als Organellen im weiteren Sinn aufgefasst werden können.[2] Begriffsgeschichte und Definitionen[Bearbeiten] Möbius’ ursprüngliche Definition von „Organula“ (1882) schloss nur organähnliche Strukturen von Einzellern ein. Wilson (1900) bezeichnete Strukturen in Zellen von Mehrzellern noch als „Organe“ oder „Zellorgane“. Nach einer heute teilweise verwendeten Definition werden nur noch membranbegrenzte Strukturen als Organellen bezeichnet. Organulum (Plural: Organula) ist die Verkleinerungsform zum lateinischen Organum. „Die Organe der Heteroplastiden (= Mehrzeller) bestehen aus vereinigten Zellen. Membranbegrenzte Organellen[Bearbeiten] Semiautonome Organellen[Bearbeiten]
Filamin Filamine (FLN) sind Proteine bei Eukaryoten und gehören zu den Aktin-bindenden Proteinen (ABP). Sie sind an der Quervernetzung von Aktinfilamenten, einem Hauptbestandteil des Zytoskeletts, sowie der Vernetzung von Aktinfilamenten mit Proteinen in der Zellmembran beteiligt. Indirekt spielen sie darüber hinaus eine wichtige Rolle bei Zell-Zell- und Zell-Matrixverbindungen, der Fortbewegung von Zellen (Zellmotilität) sowie bei der Entwicklung von Eukaryoten (Ontogenese). Mutationen in den für Filamin-B oder -C codierenden Genen können seltene Erbkrankheiten verursachen.[1] Nomenklatur[Bearbeiten] Die Nomenklatur war lange Zeit relativ uneinheitlich. hs für Homo sapiens (Mensch)gg für Gallus gallus (Bankivahuhn)dm für Drosophila melanogaster (Schwarzbäuchige Taufliege)dd für Dictyostelium discoideum (Art aus der Klasse der Schleimpilze) Eine Übersicht über die verschiedenen Filamine bietet die folgende Tabelle: [2] [3] Die Filamine des Menschen[Bearbeiten] Aufbau[Bearbeiten] M.
Ektoplasma Eine Amöbe in einer Zeichnung von 1900: Das Endoplasma füllt fast den ganzen Zellbereich aus, das Ektoplasma ist dagegen nur am Rand vorhanden. Das Ektoplasma, auch Außenplasma oder Ektosark, ist die äußere Schicht des Zytoplasmas vieler einzelliger Organismen.[1] Den innenliegenden Teil des Zytoplasmas dagegen bezeichnet man als Endoplasma, wobei beide Plasmaarten nicht grundsätzlich verschieden sind, sondern sich lediglich in ihrer Konsistenz und unterschiedlichen Verteilung intraplasmatischer Organellen voneinander unterscheiden.[2] Aufbau und Funktion[Bearbeiten] Das Ektoplasma enthält im Gegensatz zum Endoplasma keine Organellen. Vor allem bei der Ausbildung von Pseudopodien kommt es zu einer Vermischung beider Bestandteile (Gel-Sol-Übergänge); das Ektoplasma verflüssigt sich und fließt in Richtung des Pseudopodiums, während das Endoplasma in das Pseudopodium fließt und am Rand viskoser wird. Einzelnachweise[Bearbeiten]
Cyclose Un article de Wikipédia, l'encyclopédie libre. Portail de la biologie cellulaire et moléculaire Endoplasma Eine Amöbe in einer Zeichnung von 1900: Das Endoplasma füllt fast den ganzen Zellbereich aus, das Ektoplasma ist dagegen nur am Rand vorhanden. Das Endoplasma, auch Entoplasma oder Entosark, ist der zentrale, innere, Teil des Zytoplasmas vieler einzelliger Organismen.[1] Den außenliegenden Teil des Zytoplasmas bezeichnet man als Ektoplasma, beide unterscheiden sich in ihrer Konsistenz voneinander. Beide Plasmaarten sind nicht grundsätzlich verschieden sondern unterscheiden sich nur in der unterschiedlichen Verteilung von intraplasmatischen Organellen.[2] Aufbau und Funktion[Bearbeiten] Das Endoplasma enthält alle zentralen Organellen wie den Zellkern, Mitochondrien, endoplasmatisches Retikulum sowie Nahrungsvakuolen und andere Bestandteile der Zelle.[1] Im Gegensatz zum viskosen Ektoplasma (Plasmagel) hat das Endoplasma eine flüssige Konsistenz und wird daher als Plasmasol bezeichnet. Belege[Bearbeiten] ↑ Hochspringen nach: a b c Stichwort „Endoplasma“ in: Herder-Lexikon der Biologie.
Cytoplasma Als Zytoplasma, oder Cytoplasma, (von altgriechisch κύτος kýtos „Gefäß“, „Höhlung“ sowie πλάσμα plásma „Gebilde“)[1] wird die die Zelle ausfüllende Grundstruktur bezeichnet, die aus dem flüssigen Zytosol (Matrix cytoplasmatica) mit verschiedenen gelösten Stoffen sowie dem festeren Cytoskelett (Cytoskeleton) besteht. Es wird nach außen von der Zellmembran (Membrana cellularis, Cytolemma oder auch Plasmalemma bei Pflanzenzellen) begrenzt. In ihr sind Organellen (Organella) inklusive Zellkern (Nucleus) enthalten und durch verschiedene Membranen gegen das Zytoplasma abgegrenzt (Diese Sichtweise ist jedoch umstritten; einige betrachten die Organellen als Bestandteil des Zytoplasmas). Eingeführt wurde das Wort „Zytoplasma“ von dem deutschen Anatomie-Professor Albert von Kölliker 1863 als Synonym für „Protoplasma“. Begriffe[Bearbeiten] Das Zytoplasma ist die Grundsubstanz der Zelle. Funktion[Bearbeiten] Chemische Zusammensetzung des Zytoplasmas[Bearbeiten] Literatur[Bearbeiten] Plasmaströmung
Pseudopode Un article de Wikipédia, l'encyclopédie libre. Sur les autres projets Wikimedia : pseudopode, sur le Wiktionnaire C'est un prolongement cellulaire rétractile qui permet à la cellule de ramper le long d'une surface. Caractéristiques[modifier | modifier le code] On observe la production de pseudopodes uniquement chez les cellules eucaryotes, plus précisément chez les cellules animales ou chez les protistes (par exemple : les amibes). Ces déformations de la cellule sont provoquées par des pressions internes du cytoplasme. Le cytoplasme existe sous deux formes : un état liquide, la solution colloïdale (SOL) ;un état semi-solide (GEL). Le passage du cytoplasme de l'état SOL à l'état GEL facilite la formation de pseudopodes. Le cytoplasme à l'état SOL est repoussé contre la membrane plasmique située dans le sens du mouvement là où le réseau d'actine a été démonté (c'est-à-dire affaibli). Le réseau d'actine se désassemble permettant l'allongement du pseudopode. Voir aussi[modifier | modifier le code]